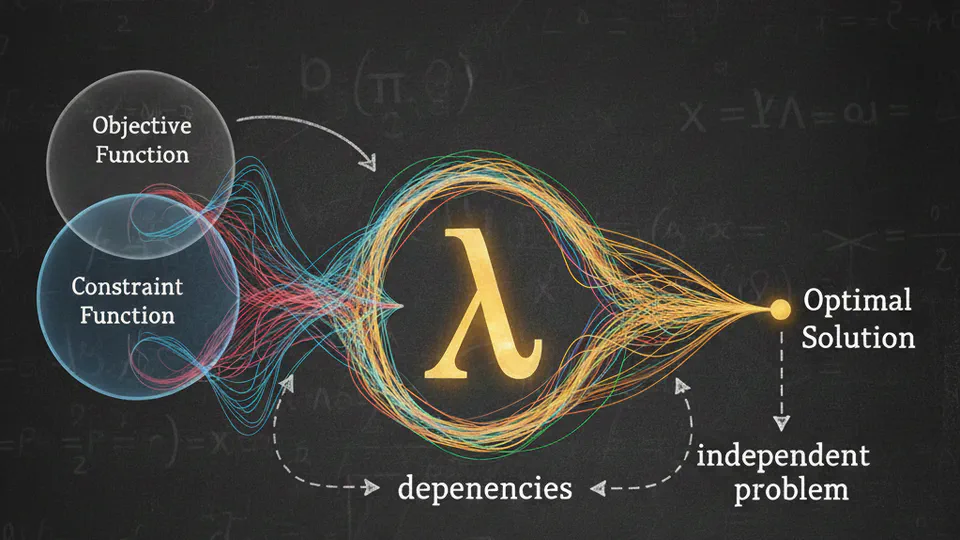
A deep dive into why we can treat dependent variables as independent when using Lagrange multipliers, and how the multiplier absorbs the constraint's effects.
A step-by-step derivation of the Poisson log-likelihood and maximum likelihood estimators for gene expression counts in single-cell data.
A comprehensive mathematical derivation of DeepSeek MLA's weight absorption mechanism, explaining how it compresses the KV cache by 57× while maintaining performance.
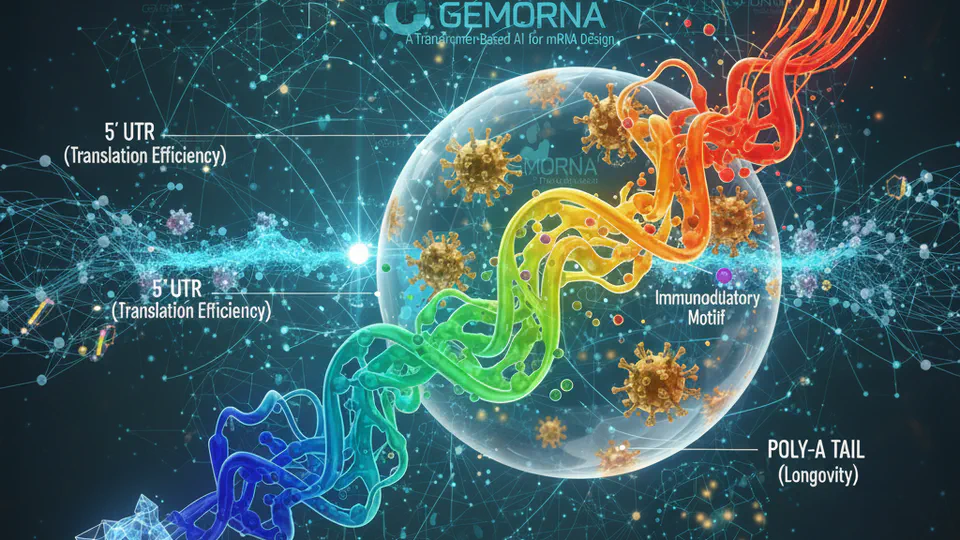
Exploring the architecture, training strategies, and surprising emergent capabilities of GEMORNA models for comprehensive mRNA design.
Stochastic model of RNA velocity defined by the Chemical Master Equation (CME) is superior to the deterministic ODE model in capturing the stochastic nature of single-cell RNA sequencing data. I derive the key equations from the paper [velocity unraveled](https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1010492) and explain their biological significance.
The steady‑state model was the first to enable a mathematical estimation of RNA velocity, and most subsequent methods are modified versions of it or its generalization (the dynamic model in `scVelo`; see our other blogs). It has its limitations and a solid understanding of its underlying mathematics is needed to apply the model effectively. Here, we derive the steady-state model in `scVelo` and `velocyto`.
Here reveals biologically significant tumor development trajectory from real-world scRNAseq dataset by tweaking the key steps and parameters in scVelo pipeline to align with the math foundations of dynamic RNA velocity model and to improve the estimation accuracy.
Here delves into the Gillespie Stochastic Simulation Algorithm to generate high-quality simulated data with known ground truth. The simulation rigorously validate the adjustments of key (hyper)parameters and processing steps in scVelo for more accurate and meaningful analysis.
Here begins the deep dive into scVelo’s computational handling that is only unveiled in its implementation but reshapes visualization and interpretation. We look into neighbor reliance, seed dependency and object structure of scVelo in this sixth blog on effectively applying the dynamic model of RNA velocity.
scVelo source codes shows a ad-hoc voting method to calculate a global latent time, which was identified as key weaknees. Here present alternative methods to potentially address the relative scale of different genes and avoid the assumption of equal full cycle time for all genes.